

서 론
미생물을 이용한 황동석 생물학적 침출
황동석의 전기화학적 특성
황동석 생물학적 침출 전기화학적 원리와 미생물의 역할
생물학적 침출의 미생물 특성
용액전위(Fe3+/Fe2+ 농도비)에 따른 황화광 침출 조절
용액전위(또는 Fe3+/Fe2+농도비)에 따른 황동석 침출조절
해결해야 할 문제
결 론
서 론
생물학적 침출은 미생물을 이용하여 광석내 유용금속을 침출시키는 기술로 저품위 금속회수에 유리하고 환경친화적인 기술로 인식되고 있다(Debernardi and Carlesi, 2013; Panda et al., 2015; Zhao et al., 2015). 특히 Cu 선광의 경우 최근 환경오염의 심각성과 Cu광체의 저품위화로 인하여 생물학적 침출의 상용화가 활발하다(Panda et al., 2015; Schippers et al., 2014). 실제 생물학적 침출을 이용한 Cu 회수는 1950년도에 상업화되었다(Panda et al., 2015). 다만 현재 Cu 부존량의 대부분을 차지하는 황동석 선광의 경우에는 내화학성이 높고 침출반응 중 부동태층의 생성으로 인해 생물학적 침출이 아직 상용화되지 못하고 있다(Debernardi and Carlesi, 2013) 그럼에도 불구하고 최근 전세계적으로 Cu광체의 저품위화가 심화되고 있어 경제성 있는 황동석 생물학적 침출 기술개발이 절실한 실정이다(Feng et al., 2014).
Cu 생물학적 침출 기술은 Heap/Dump 침출 또는 교반탱크를 이용한 생물학적 침출 등으로 구분되는데 이중 실제 상용화된 것은 대부분 2차 Cu황화광을 대상으로 한 Heap/Dump 생물학적 침출 기술이다(Clark et al., 2006; Panda et al., 2015; Schippers et al., 2014; Zou et al., 2015). 최근 생산된 전체 Cu량의 7% 정도가 Heap 생물학적 침출을 거쳐 생산되었으며 2014년 세계적인 구리 생산국인 칠레와 페루의 경우 각각 42%, 33%의 Cu를 생물학적 침출을 통해 생산하고 있다(Panda et al., 2015). 전세계적으로 저품위 황화광내 Cu 개발 필요성이 대두됨에 따라 생물학적 침출의 상용화는 확대될 예정이다. 다만 2차 Cu 광물선광이 생물학적 침출이 활발하게 적용되는 반면 황동석의 생물학적 침출은 주로 파일롯 혹은 소규모 시범 플랜트로 운영 중이다(Panda et al., 2012; Panda et al., 2015).
Fig. 1은 가장 일반적인 형태인 Heap 생물학적 침출을 묘사한 것이다. 비교적 균일한 크기의 Cu 광석을 쌓은 Heap내에 호기성/호산성 철/황산화 미생물을 접종한 후 Heap 상부에서 미생물, 영양분, pH 조절제, 환원제 등을 포함한 수용액을 뿌리고 하부에서는 공기를 불어넣어 호기성 미생호기성 미생물에 의해 금속이 침출되도록 한다. Heap 하부에 모인 침출수는 집수한 후 하부 공정인 SX-EW(Solvent Extraction & Electro-winning) 등의 공정을 거쳐 고순도 Cu를 회수한다. 침출수가 토양 및 지하수로 빠져나가지 못하도록 Heap 하부에 Leach pad를 설치한다(Lupo, 2010). 또한 집수된 침출수는 다시 Heap 상부로 올려보내 재사용함으로써 액중 Cu농도가 적절한 수준에 이르게 한다. 강우량이 많은 지역에서는 비가 Heap 내부로 흘러들어가지 않도록 Heap 상부에 덮개를 씌우는 경우도 있다(Panda et al., 2015).
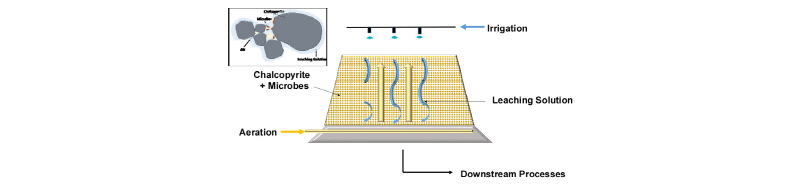
미생물을 이용한 황동석 생물학적 침출
미생물이 생존하고 증식하기 위해서는 탄소원, 에너지원 그리고 영양분을 필요로 한다. 이러한 대사화학반응을 간단하게 기술하면 다음 식 (1)과 같다(Hatzikioseyian and Tsezos, 2006; Rittmann and McCarty, 2001).
탄소원 + 에너지원 + 최종 전자 수용체 + 영양분
→ 미생물체 + 대사산물 (1)
퇴비화 또는 일반하수처리 같이 호기성 생분해에 사용되는 미생물(유기종속영양균)은 주로 유기물을 탄소원 및 전자공여체로 이용하고 용존산소 O2를 최종전자수용체(호흡)로 하여 증식 및 생존에 필요한 에너지를 얻는다. 반면, 생물학적 침출에 주로 참여하는 호기성 미생물(무기독립영양균)은 주로 CO2를 탄소원으로 하고 별도로 Fe2+과 S을 전자공여체로 하며, O2를 최종 전자수용체로 하여 에너지를 얻는다. 황동석 생물학적 침출은 미생물이 광물내 주로 Fe을 산화시켜 증식과 성장에 이용하고(S도 이용하나 Fe 산화에 비하면 에너지가 적다) 남은 Cu는 화학적으로 침출되는 것이다. 무기독립영양 미생물이 얻을 수 있는 에너지나 증식량은 유기종속영양균에 비하면 많지 않은 편이다. 그래도 증식에 필요한 약간의 탄소원인 CO2 외 에너지원인 Fe/S/Cu와 최종 전자 수용체인 O2그리고 별도의 영양분이 적절히 주입되어야 한다(Rittmann and McCarty, 2001).
황동석 생물학적 침출에 관련된 메커니즘은 다양하고 복잡하며 여전히 논란이 많다. 심지어는 미생물이 금속침출에 어떤 영향을 미치는지, 구체적으로 어떠한 반응경로 및 에너지로 황동석내 Cu가 침출되고 저해되는지도 명확하게 규명되지 않았다(Debernardi and Carlesi, 2013). 이러한 생물학적 침출을 분석하고 이해하는데는 열역학적 관점(Hatzikioseyian and Tsezos, 2006), 반응속도(Ojumu et al., 2006) 및 공학적 모델링(Petersen, 2010) 관점 등 여러 가지 기법이 있으나 이중 전기화학적 개념을 이용하는 것이 가장 효과적이고 중요한 것으로 인식되고 있다(Debernardi and Carlesi, 2013; Khoshkhoo et al., 2014; Vargas et al., 2014). 따라서 본 연구에서는 이러한 추세에 따라 황동석의 생물학적 침출을 주로 전기화학적 관점에서 논의하고자 한다.
황동석의 전기화학적 특성
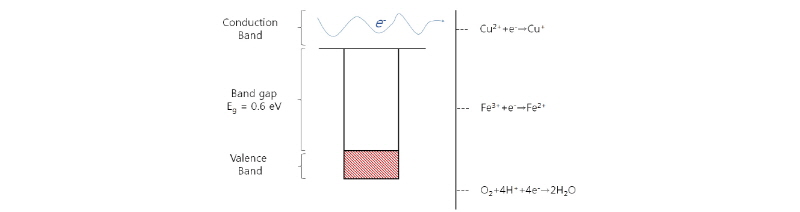
황동석(밀도 대략 4.2 g/cm3)는 간단한 화학식으로 CuFeS2로 표기하는데 황원자가 Cu 또는 Fe 금속원자와 결합되어 있는 물질이다(Debernardi and Carlesi, 2013). Lattice energy가 17500 kJ 정도 되는데 이는 여타 황철석, Covellite, 휘동석의 3000-4000 kJ에 비해서 훨씬 높은 값이다. 즉 다른 황화광보다 내화학성이 높아 침출시키기 어렵다(Debernardi and Carlesi, 2013). 또한 황동석 물성에서 반드시 언급되어야 할 점은 황동석이 천연 반도체 물질이라는 사실이다(Crundwell, 1988). 많은 전도성 금속화합물의 경우 전도대(Conduction band)와 가전자대(Valence band)가 겹쳐있어 가전자대내 전자가 전도대로 쉽게 이동, 자유전자의 형태로 돌아다니는 반면, 황동석 같은 반도체 물질에서는 그런 전자의 움직임이 보다 제한된다(Crundwell, 1988; Crundwell, 2015; Debernardi and Carlesi, 2013; Li et al., 2013). 그 이유는 (1) 결합된 전자가 위치한 가전자대 (2) 결합되지 않은 자유전자가 돌아다닐 수 있는 전도대 그리고 두 밴드(Band)사이의 밴드갭(Band gap)이라는 일종의 에너지 간극이 존재하는데 이 간극 내에는 전자가 존재할 수가 없다. 황동석 내 Fe 전자의 경우 Conduction band의 반결합궤도(Anti-bonding orbital)에 주로 전자가 있고 반대로 Cu의 전자 결합궤도는 가전자대에 있다. 즉 결정 내에서 Cu는 강하게 S과 결합되어 있다. 전도대와 가전자대 사이의 갭은 대략 0.6 eV이다(Crundwell, 1988; Crundwell, 2015; Debernardi and Carlesi, 2013; Vargas et al., 2014). 황동석의 전자 밴드 diagram을 Fig. 2에 나타내었으며, 주요 산화/환원 전위값을 그림 오른편에 정렬하였다.
황동석 생물학적 침출 전기화학적 원리와 미생물의 역할
Fig. 2를 보면 Cu2+/Cu+ 표준환원전위(E0 = 0.16V), Fe3+/Fe2+의 표준환원전위 (E0= 0.77V) 및 O2/H2O의 표준환원전위(E0 = 1.23V)순으로 환원성이 강하다고 볼 수 있다. 또한 황동석 내 Fe의 자유전자는 전도대 준위에 있기 때문에 Cu보다 쉽게 산화/침출될 수 있다. 호기성 미생물에서 자주 나타나는 O2 + 4H+ + 4e- → H2O의 환원반응의 Coupling으로 Fe2+는 다시 산화되며, 이 때 발생한 전자가 산소의 환원에 이용된다. Fig. 3은 호기성 미생물의 대사로 인해 발생하는 일련의 연쇄반응 즉 Fe3+/Fe2+의 산화/환원 및 황화광 용출 반응을 도식화하였다.
|
Fig. 3. Fe3+/Fe2+ oxidation and reduction and consequential bioleaching of metal sulfide (MeS) by aerobic chemolithotrophs. Adapted from Vargas et al. (2014). |
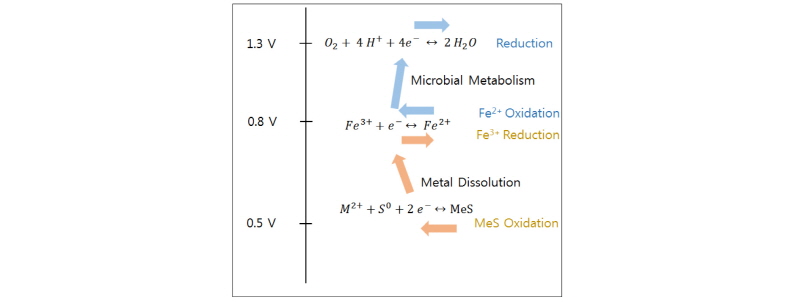
생물학적 침출은 황화광(MeS)내 금속을 침출시키는데, 특히 황동석 생물학적 침출의 경우 Fe2+과 S를 산화시킬 수 있는 중/고온성 미생물을 이용하여 Cu를 침출시킨다. 이 경우 철이 이온상태로 안정하게 존재하려면 강산성이어야 하기 때문에 생물학적 침출내 철산화 미생물은 그러한 강산성 환경에 견딜 수 있어야 한다(또한 호기성 조건하 산소환원 및 침출반응자체의 증진을 위해서도 많은 H+이온이 필요하다). 황동석의 침출반응 관련해서는 다양한 견해가 있으나(Debernardi and Carlesi, 2013), 기존 문헌에서 가장 자주 인용되는 반응식(Dutrizac, 1981; Hackl et al., 1995; Hiroyoshi et al., 2002; Yoo et al., 2008) 을 기술하면 다음 식 (2)와 같다.
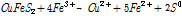 (2)
(2)
상기 식을 보면 황동석내 Fe, Cu이 산화침출되고 S도 S0(원소황)으로 산화되었다. 이 산화반응이 Fe3+의 환원반응과 Coupling을 이루고 있다. 그런데 별도의 Fe3+ 공급 없이 침출반응이 진행되다 보면 Fe3+가 계속 소모되어 결국 고갈될 수 밖에 없게 된다. 이 경우 식 (3) 같은 미생물 반응을 통해 침출반응에 필요한 에너지(전위)와 Fe3+를 재생/공급한다(Debernardi and Carlesi, 2013; Vargas et al., 2014).
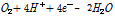
 (3)
(3)
위 식 (3)에서 알 수 있듯이 황동석의 생물학적 침출에서 미생물의 가장 중요한 역할 중 하나는 식 (2)의 생성물인 Fe2+을 산화시켜 Fe3+을 재생시킴으로써 침출반응을 계속 유지하는 것이다(Li et al., 2013; Zeng et al., 2010). 이때 Fe2+의 산화반응 결과 생성된 전자는 높은 환원전위를 가진 O2의 환원반응에 이용된다. 즉 미생물에 의해서 O2가 물로 환원되면서 생성된 높은 에너지로 Fe2+를 산화시켜 Fe3+을 생성되고 황동석내 Cu와 황을 침출시킬 수 있다는 의미이다(Vargas et al., 2014). 이러한 반응물이 미생물체내 혹은 부근에 농집되어 있기 때문에 광석표면의 작은 금속입자도 추출이 가능한 것이다. 또 다른 미생물의 역할로는 상기 침출의 또 다른 생성물인 S0를 산화시킨다(Debernardi and Carlesi, 2013; Li et al., 2013). 미생물은 S0의 표면축적을 방지하고 동시에 pH를 낮추어 침출반응을 지속적으로 활성화시킬 수 있다. 이는 다음 식 (4)에 의해 나타낼 수 있다.
 (4)
(4)
생물학적 침출의 미생물 특성
종래에는 생물학적 침출에 참여하는 미생물 형태를 Fe을 산화시키는 반응특성에 따라 간접, 직접 메커니즘으로 구분하였으나(Rohwerder et al., 2003) 최근에는 미생물의 광석표면 부착여부에 따라서 접촉, 비접촉, 협력적 메카니즘으로 세분화하였다(Watling, 2006).
간접적 메커니즘은 미생물이 용액 내에서 Fe2+를 Fe3+로 산화시키는 반응을 의미한다. 한편 직접적 메커니즘은 이러한 Fe3+/Fe2+산화과정 없이 미생물이 직접 황화물을 산화시키는 반응을 의미한다. 이 중 접촉/비접촉에 상관없이 간접적 메커니즘이 잘 규명되어 있는데 본 연구에서 지금까지 기술한 Fe2+의 산화를 통한 Fe3+ 재생산이 모두 간접적 메커니즘의 결과이다. 즉 간접 메카니즘은 본질적으로 화학적 침출의 그것과 거의 같으나 미생물의 Fe2+ 산화를 통해 화학적 침출반응을 계속 유지시킨다는 점만 다르다. 반면 직접 메커니즘에 대해서는 실험적으로 검증된 바가 없다(Panda et al., 2015; Shippers et al., 2014).
한편 접촉 메커니즘은 광석표면에 부착된 미생물의 생촉매반응(Enzymatic reaction)에 의해 Fe2+가 산화된다(Rodríguez et al., 2003). 이때 미생물의 부착은 초기에는 주로 정전기적 또는 기타 물리적(미생물 표면의 섬모) 작용을 통해 가역적으로 광석표면에 부착하지만 시간이 지나면서 미생물 군집을 형성하게 되고 EPS(Extracellular Polymeric Substance) 형성을 통해 미생물 군집이 표면에 보다 견고하게 부착된다(Rodríguez et al., 2003). EPS는 미생물에서 배출된 일종의 점액분비물로 미생물 군집을 외부 환경으로부터 보호하여 미생물 생태계의 안정성을 증대시키는 역할을 한다. 또한 EPS를 통해 Fe2+이나 황화물을 산화시키기도 한다(Li et al., 2013). Fe2+나 황화물의 산화는 EPS가 없는 미생물계에서도 가능하지만 EPS는 반응관련 물질을 체내에 농축시킬 수 있어 광석내 작은 금속조각도 효과적으로 침출시킬 수 있다(Kinzlera et al., 2003; Sand and Gehrke, 2006). 즉 EPS가 생물학적 침출의 저품위 광물 침출의 고효율화에 큰 역할을 하는 것으로 보이지만 한편으로는 EPS의 과다 생성이 광석의 침출을 저해하기도 한다(Zeng et al., 2010; Zeng et al., 2011). 부착 생물막의 증식은 용액환경(유체흐름, 온도, pH 등)외에도 부착대상인 광석표면의 접촉면적, 공극구조, 결합에너지, 미생물 친화성(표면전위에 따른 정전기적 인력 혹은 척력 등)에 영향을 크게 받는다(Li et al., 2013; Rodríguez et al., 2003). 비접촉메커니즘은 표현 그대로 부유 미생물(Suspended cell)에 의해 Fe2+가 산화되는 반응이다(Li et al., 2013; Watling, 2006). 마지막으로 협력적 메커니즘은 생물막이 포화되었을 때 접촉 미생물과 부유미생물의 일종의 공생계를 이루는 것이다(Gautier et al., 2008; Li et al., 2013; Watling, 2006). 이는 생물학적 침출 효율을 높이는 역할을 한다. Fig. 4는 미생물 반응형태에 대해서 묘사한 것이다.

미생물종 측면에서는 특히 고온성 미생물이나 혼합 미생물을 투입하였을 때 생물학적 침출효과가 증가하는데, 전자는 가용에너지가 크고 후자의 경우 생태적 안정성의 측면에서 단일종보다 혼합종이 적응 및 증식에 유리하기 때문인 것으로 추측된다(Ahmadi et al., 2011; D’Hugues et al., 2002; Zeng et al., 2010; Zeng et al., 2011). 특히 고온성 미생물을 아주 미세하게 분쇄된 광석과 반응시킬 때 침출효과가 매우 높다 . 반면 중온성 미생물은 고온성 미생물에 비해 생물학적 침출 효과가 낮은 것으로 나타났다(Olson etal., 2003).
실제 운전측면에서 생물학적 침출이 화학적 침출과 크게 다른 점 중 하나는 미생물이 반응계에 충분히 적응하고 미생물량이 적절히 확보될 때까지 상당한 적응기간이 필요하다는 점이다. 이로 인해 사전 운전기간이 길어질 수 있다. 미생물의 생존조건으로는 일반적으로 적정 습도(수분함유량), pH, 온도, 낮은 독생/저해물질 및 충분한 에너지원과 영양물질 등이 중요한 것으로 알려져 있다(Li et al., 2013; Panda et al., 2015; Vilcáez et al., 2008; Watling et al., 2006). 특히 생물학적 침출에 자주 사용되는 무기독립영양균의 경우 이화작용(호흡 등을 통해 에너지를 얻는 대사과정)이 무기질(Fe이나 S)의 산화에 많이 의존하기 때문에 이들의 산화/환원전위가 매우 중요하다(Rittmann and McCarty, 2001). 생물학적 침출의 경우 여기에 반응용액의 산화환원전위, 예를 들면 Fe3+/Fe2+농도비가 적절해야 한다. 또한 광석의 크기는 작을수록 비표면적이 증가하여 유리하다.
또한 황동석 생물학적 침출 관련 추가적인 주요인자는 유사 광물 침출시 대비 약간 낮은 전위, 고온, 그리고 적정한 양의 촉매(Ag) 등이 있다. 이 중 Fe3+와 Fe2+의 농도비 및 전위가 특히 중요하다(Vargas et al., 2014). 생물학적 침출은 대부분 산화반응으로 Coupling이 될 환원반응(Fe3+ → Fe2+)에 많은 Fe3+가 소모되며 또한 환원된 Fe2+는 호산성 철산화 미생물의 에너지원으로 쓰이기 때문에 역시 충분한 양의 Fe2+도 필요할 것이다. 여기서 중요한 것은 Fe3+/Fe2+ 농도비인데 이 비율이 사실상 용액전위를 결정한다(다른 Cu2+ 이온 등의 영향을 무시한다는 가정 하에). 전기화학적 고전위는 Fe2+ 대비 고농도의 Fe3+로 인해 산화침출이 활성화된다는 의미이고 반대로 저전위는 Fe3+ 대비 고농도의 Fe2+로 인해 미생물의 철산화 반응이 활발하게 발생할 수 있다는 의미이다. 이 같이 Fe2+/Fe3+ 농도를 이용한 황화광 생물학적 침출 반응속도 조절의 개념을 다음 장에 보다 상세히 논의하였다.
용액전위(Fe3+/Fe2+ 농도비)에 따른 황화광 침출 조절
본 장에서는 황동석 생물학적 침출에 앞서 일반적인 황화광(MeS ↔ Me2+ + S0 + 2e-)을 대상으로한 생물학적 침출반응속도가 어떻게 Fe3+/Fe2+농도비에 의해서 조절되는지 논의한다. 전기화학적 반응예측에서는 전위차를 이용하여 각 반응의 자발성 여부를 판단하는 것이 가장 중요하지만 각 반응의 속도도 중요한 고려사항이다. 문제는 반응속도론 관점에서 기존의 생물학적 침출의 반응속도의 모델화 관련 연구가 충분치 않다는 점이다(Ojumu et al., 2006). 기존 연구 중 가장 간단한 반응속도식을 예로 설명하자면, Vargas et al. (2014)의 경우 전기화학적 전위를 기반으로 매우 간단한 반응속도 모델을 개발하였는데 여기서 중요한 가정은 임의의 두 반응간(R1, R2) 전자이동속도는 두 반응의 전기화학적 전위차(ER1-ER2)와 밀접한 상관관계가 있다고 본다는 것이다. 즉 ER1-ER2가 증가할수록 전자이동속도 즉 반응속도는 증가한다고 전제한다. 그러면 반응평형에 포함되었던 Fe3+/Fe2+의 농도비가 생물학적 침출 속도식에도 당연히 중요한 인자로 포함된다. 이 때 다른 조건 즉 Me2+, H+농도 그리고 산소분압 P(O2)이 일정하다는 다른 가정이 필요하다. Fe3+/Fe2+ 농도비가 증가할 경우 황동석의 산화용출 반응속도가 증가할 것이며 반대로 농도비가 감소할 경우 산화반응속도는 감소할 것이다(Vargas et al., 2014). Fe3+/Fe2+의 비는 다음과 같이 전기화학적 전위로 나타낼 수 있다.
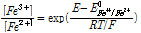 (5)
(5)
여기서 E는 용액의 환원전위(V), E0Fe3+/Fe2+는 Fe3+의 표준환원전위(V) R은 기체상수, T는 절대온도(K), F는 Faraday 상수이다(C/mol). 식 (5)를 기반으로 미생물에 의한 Fe2+의 산화반응 속도식을 다음과 같이 표현할 수 있다(Boon et al., 1998; Breed et al., 1999).
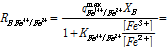
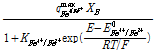 (6)
(6)
식 (6)은 Fe3+/Fe2+ 농도비를 저해인자로 상정한 일종의 Michaelis-Menten 형태의 속도식((Jones and Kelly, 1983)으로 미생물에 의한 Fe2+산화속도를 RB_Fe3+/Fe2+로 나타내었다. 여기서 qmaxFe3+/Fe2+는 최대 미생물 산화속도(1/s)를 의미하고 XB는 반응에 참여하는 활성화 미생물의 농도(mg/L), 그리고 KFe3+/Fe2+는 본 미생물 산화속도식의 저해계수(Inhibition constant)(-)를 의미한다.
한편 아래 식 (7)은 침출시의 Fe3+의 환원속도(혹은 황화광 MeS의 침출속도) RC_fe3+/Fe2+를 나타낸 것이다.
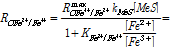
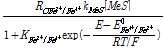 (7)
(7)
여기서 RmaxC_Fe3+/Fe2+는 최대 침출반응속도(1/s), kMeS는 황화광 표면반응속도관련 계수(-), 그리고 KFe2+/Fe3+는 본 침출반응속도식의 저해계수(-)이다.
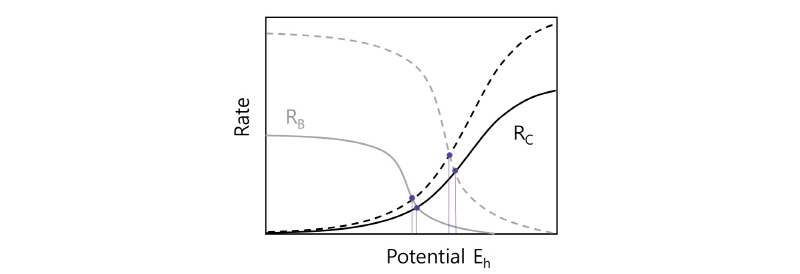
상기 식 (6), (7)을 구현한 속도곡선을 Fig. 5의 그래프에 나타내었다(Vargas et al., 2014). 그래프에서 세로축은 미생물에 위한 산화반응속도 혹은 화학적 환원속도를, 가로축은 용액의 환원전위 Eh를 나타낸다. 가로축의 오른편으로 갈수록 환원전위가(혹은 Fe3+/Fe2+의 농도비가) 증가한다. 두 곡선 중 왼쪽 곡선의 형태를 보면 Eh가 감소할수록(가로축의 왼쪽으로 갈수록) RB_Fe3+/Fe2+는 급격히 증가하는데 Eh가 계속 감소하게 되면 더 이상의 증가가 거의 없는 일종의 평탄선(Plateau)에 도달한다. 이는 미생물 활동이(미생물의 증가가 없다는 가정 하에) 최대 산화율 qmaxFe3+/Fe2+에 도달했다는 것을 의미한다(Vargas et al., 2014).
|
Fig. 5. The curves representing the microbial oxidation rates of Fe2+ (RB) and the reduction rates of Fe3+ (RC) due to anodic dissolution of chalcopyrite. Modified from Vargas et al. (2014). |

반면 화학적 반응속도(오른쪽) 곡선에 나타난 RC_Fe3+/Fe2+은 Eh가 증가할수록 처음에는 가파르게 증가하다가(침출속도가 빨라지다가) 더 이상 증가하지 않고 어떤 평탄선에 이르게 된다. 이 두 속도 곡선이 교차하는 지점은 Eh이 대략 850 mV가 되는 지점으로 두 반응 즉 MeS의 화학적 침출속도(식 (7))와 Fe2+의 전자가 O2로 전이되는 미생물 산화반응속도(식 (6))가 일치하는 지점이다. 이 지점에서 Fe3+/Fe2+가 더 이상 변하지 않는다고 보고 있다(단 여기서 kMeS[MeS]나 미생물량 XB가 일정하다고 가정한다). Eh가 정상상태 즉 850 mV에 이르기 전까지 Fe3+/Fe2+의 비율은 다음 속도식에 따라 계속 변한다고 본다. 예를 들면 Eh>850 이면 Fe2+는 증가하는(용액전위 E가 낮아지는) 반면 Eh<850 이면 Fe2+는 감소한다. 용액전위 Eh 또는 Fe3+/Fe2+를 통한 Fe2+ 농도변화율은 다음과 같다.
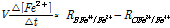 (8)
(8)
여기서 V는 시스템의 부피(L) 그리고 t는 반응시간(s)을 의미한다. 시간에 따라서 식(8)의 좌우항은 모두 0에 수렴하게 된다 (Fe2+의 농도가 정상상태에 이른다).
식(6), (7)을 다시 보면 실제로 시간에 따라 전위 Eh만 변하는 것이 아니라 미생물량 XB 또는 침출대상인 황화광 표면 반응지점[MeS]도 변화하게 된다. 따라서 시간에 따라 산화곡선 자체도 변화하게 되는데 미생물량은 대체로 시간에 따라 증가하게 되므로 미생물 산화곡선은 상승하게 된다. 반면 [MeS]는 시간에 따라 감소하고 특히 표면에 부동태층이 축적될 경우 크게 감소하게 된다(Debernardi and Carlesi, 2013). 이 경우 정상상태에 수렴되는 최종전위는 크게 변화한다. Fig. 6은 생물학적 또는 화학적 조건 변화에 따른 최종 전위의 변화를 나타낸 것이다.
용액전위(또는 Fe3+/Fe2+농도비)에 따른 황동석 침출조절
식 (6)-(8)을 이용한 반응속도 모델은 어디까지나 일반적인 황화광 생물침출을 대상으로 한 개념적 모델로서 실제 황동석의 침출과는 많은 차이를 보인다. 대표적으로 Hiroyoshi et al. (2001)은 황동석 화학적 침출실험 결과 저전위하(560–600 mV)에서 Fe3+ 외에도 고농도 Fe2+와 저농도 Cu2+조건에서 황동석의 산화침출속도가 증가한다는 사실을 발견했다. 이는 상기 식 (6)-(8)과는 맞지 않은 결과였다(Nicol et al., 2010; Hiroyoshi et al., 2004; Hiroyoshi et al., 2008). Hiroyoshi et al. (2008)는 이 현상을 근거로 새로이 2단 모델(Two step model)을 제안하였다. 이 모델에 따르면 1단계에서 황동석이 Cu2+이온의 존재 하에 Fe2+에 의해 보다 침출이 용이한 휘동석으로 환원되고, 2단계에서 휘동석이 산화되면서 Fe3+의 환원을 수반한다. 이를 반응식으로 표현하면 다음과 같다.
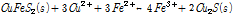 (9A)
(9A)
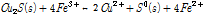 (9B)
(9B)
여기서 식 (9B)에 2를 곱하고 식 (9A)를 더하면 식 (2)가 산출된다. 즉 Hiroyoshi et al. (2008)의 반응식은 기존 반응식(식 (2))을 2단계로 세분화한 것이다.
본 모델에 따르면, 이 두 반응이 모두 자발적으로 발생하려면 전위 Eh가 (1) 1단계 반응에서 황동석이 휘동석으로 환원될 수 있을 만큼 낮고, (2) 2단계 반응에서 휘동석이 충분히 산화될 수 있을 만큼 높아야 된다. 이는 (1)과 (2) 모두를 만족시키는 최적 산화/환원 전위가 존재한다는 것이다. 최적 Eh값은 조건에 따라 다양하지만 일반적으로 630 mV (vs Ag/AgCl) 정도로 알려져 있으며 이때 액중 Cu의 농도는 50-200 mg/L 범위 내여야 한다고 보고하였다. (Debernardi and Carlesi, 2013; Hiroyoshi et al., 2008).
식 (8)의 반응속도식에 Hiroyoshi et al. (2008)이 제시한 개념을 고려하면 생물학적 침출 반응속도식은 더 복잡한 양상을 띠는데 아직 미생물 대사까지 고려한 반응속도 측면에서는 일반적인 황화광 MeS만 고려한 모델보다 연구가 충분하지 않다. 따라서 본 연구에서는 이를 기존 문헌의 내용을 가지고 간략하게만 논의한다. 우선 Hiroyoshi et al. (2008)의 반응을 도식화하면 Fig. 7로 나타낼 수 있다.
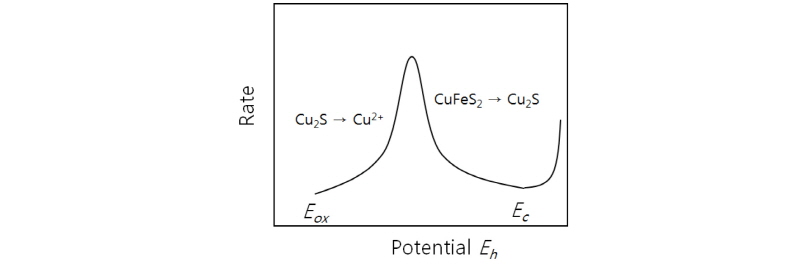
Fig. 7은 본 모델에서 산화환원전위(가로축)에 따른 황동석의 전반적 침출반응속도의 로그값(세로축)을 묘사한 것이다. 가로축의 변수 Eh는 Fig. 5의 경우와 마찬가지로 Fe3+/Fe2+농도비로 조절할 수 있다고 전제한다. 또한 곡선을 보면 반응식에 따라 단계별로 구분할 수 있는데 가로축의 Eh = Ec는 미생물 산화시의 전위가 황동석이 휘동석으로 환원되는데 요구되는 전위값과 같다는 것을 의미한다(식(9A)에 의한 1 단계 반응에 필요한 전위). 또한 Eh = Eox는미생물 환원전위가 1단계 반응에서 생성된 휘동석이 산화침출 반응(2단계)에 필요한 전위와 동일함을 나타낸다. 세로축을 기준으로 보면 Ec와 Eox가 낮은 반응속도를 보이는데 이는 Eh = Ec의 경우 환원반응이 우세하여 (Fe2+ 농도가 낮은 경우) 1단계 반응은 빠르게 진행되지만 2단계 산화반응이 늦어 전반적인 반응속도가 낮은 경우이고 Eox의 경우는 그 반대의 경우이다. 반응속도가 가장 높은 최적 Eh = Eop에서는 (1)+(2) 단계가 합친 전반적 침출반응속도는 가장 빠르다. 다시 말하면 황동석의 침출반응이 빨리 진행되기 위해서는 황동석이 휘동석으로 환원될 수 있을 만큼 낮아야 하며 2단계 반응에서 휘동석이 충분히 산화될 수 있을 만큼 높아야 된다. 이 두 지점을 적절히 충족시키는 전위가 Eop인 것이다(Hiroyoshi et al., 2008).
Fig. 7을 3영역으로 구분하였는데 이를 정리하면 다음과 같다.
영역 1: Eh < Eox로 전위가 낮아 휘동석 산화가 불가하여 Cu 침출이 종료된다.
영역 2: Eox <Eh < Ec로 1-2단계반응 즉 황동석의 환원과 휘동석 산화가 동시에 원활하다.
영역 3: Ec < Eh로 황동석의 환원 즉 휘동석 생성이 안되고 바로 Cu의 산화침출이 발생한다.
해결해야 할 문제
생물학적 침출은 분명 친환경적이고 저렴하며 저품위 광석에 적합한 기술이다. 이는 전세계적으로 특히 Cu와 금의 경우 품위저하가 심각하여 생물학적 침출이 좋은 대안으로 떠오르고 있다. 그러나 생물학적 침출이 황동석에 상용화되기 위해서는 먼저 다음과 같은 문제가 해결되어야 한다.
1.화학반응 측면에서 기본적인 메커니즘이 확실히 규명되어야 한다. 첫째 황동석에서 Cu 및 Fe의 침출과 황화물의 반응은 다양하고 복잡하며 이를 전기화학 및 반응속도 측면에서 규명되어야 한다. 예를 들면 Fe 이온 외에도, 저전위에서 식 (9)를 고려하여 Cu 이온농도가 포함된 속도식이 개발되어야 하며 표면 내 비균일 반응속도모델이 개발되어야 한다. 이는 여타 비산화적 또는 환원침출반응과의 반응조건과도 명확히 구분되어야 한다(Debernardi and Carlesi, 2013). 또한 반응을 검증할 관찰도구가 개발되어야 하며 특히 분광학적 미시적 관찰이 가능해야 한다.
2.미생물학적 측면에서도 확실히 규명되지 않은 메커니즘이 많은데 특히 고온성 미생물 관련 접촉, 간접, 협력적 반응 메커니즘이 더 명확히 규명되어야 한다.
3.공학적 측면에서 황동석의 Cu 침출속도 및 회수율 향상을 목적으로 하는 연구 및 기술을 증진시켜야 한다. 기존의 전위조절을 통한 성능향상 외에도 은 같은 촉매나 활성탄 같은 물질을 첨가제로 이용시 전기화학적 메커니즘 및 최적 조절 방안이 규명되어야 한다(Li et al., 2013). 또한 거시적인 기술적 문제에서도 Heap 생물학적 침출을 예로 들면 침출반응계 관련 인자 pH, 산소, 온도 구배 등를 파악하고 모델화하여 설계 및 운영상의 최적화를 도모하여야 한다.
결 론
황동석은 반도체 물질로 Cu 의 전자가 주로 결합력이 강한 가전자대내에 위치하여 Cu 산화 및 침출에 많은 에너지를 필요로 한다. 이러한 산화침출을 진행시키기 위해서는 Fe3+를 산화제로 이용하는 것이 효과적이다. 호기성 미생물에 의한 산소분자의 환원은 Fe2+를 산화시킴으로써 얻은 전자를 사용하는데 이를 통해 산화침출에 소비된 Fe3+를 재생시킬 수 있다. 또한 철이 계속 이온상태로 존재하여야 하므로 침출에 이용되는 철산화 미생물은 호산성이어야 한다. 황화광의 산화침출반응은 많은 Fe3+가 소비되어야 하므로 고전위 즉 높은 Fe3+/Fe2+ 농도비의 용액조건에서 원활하게 진행된다. 이에 반해 미생물 대사는 Fe2+의 산화를 필요로 하므로 상대적으로 저준위 즉 낮은 Fe3+/Fe2+ 농도비가 더 유리하다. 원칙적으로 황동석을 비롯한 황화광의 효과적인 생물학적 침출을 위해서는 전기화학적 측면에서 산화침출과 미생물의 산화반응이 둘다 원활하게 진행되기 위한 최적 전위 또는 최적 Fe3+/Fe2+ 농도비를 결정하는 것이 중요하다. 실제 황동석 생물학적 침출 반응은 더 복잡하다. 가장 대표적인 반응은 비교적 저전위에서 발생하는 것으로 황동석의 Cu가 바로 산화침출되는 것이 아니라 휘동석를 중간매개체로 하는 2단계 환원-산화반응이다. 첫 번째 환원 반응은 Cu가 휘동석으로 환원되는 반응이고 이 후 두 번째 반응은 휘동석에서 Cu가 침출되는 반응이다. 이 경우 황동석 Cu의 효과적인 산화침출을 위해서는 우선 첫 번째와 두 번째 반응을 모두 고려한 산화침출 최적 전위를 결정해야 하며 여기에 미생물 Fe2+의 산화를 고려하여 황동석 생물학적 침출 최적전위를 결정하여야 한다.
